
高角羚 又称黑斑羚、飞羚等,是高角羚属的唯一物种(这里补充一下,在众多记录片和书籍资料中,黑斑羚这个中文名比较常见。出于它为高角羚属中的物种,本文将以高角羚为中文名。),分布于非洲东部和南部。多见于林地,有时也出现在林地与热带稀树草原的过渡带,并倾向于栖息于近水源的区域。以日行性为主,其社会行为因气候和地理环境的不同而表现出群居性或领域性。高角羚的食谱包括各种草、嫩叶和果实。
本文基于现有文献资料,对其分类、形态、生态、行为、繁殖及保护现状进行系统综述。
界: 动物界 Animalia
门: 脊索动物门 Chordata
纲: 哺乳纲 Mammalia
目: 偶蹄目 Artiodactyla
科: 牛科 Bovidae
属: 高角羚属 Aepyceros
种: 高角羚 A. melampus

高角羚的属名Aepyceros,源于古希腊语,意为“高角”,种加词“melampus”意为“黑足”。
其最早记录的英文名称“palla”或“pallah”(1802年)源自茨瓦纳语“phala”,意为红色羚羊。“impala”这一拼写变体最早于1875年记录,直接来自祖鲁语。“rooibok”是南非荷兰语名称,有时也在英语中使用,意为红色雄兽。

高角羚由德国动物学家辛里奇·利希滕斯坦(Hinrich Lichtenstein)于1812年命名,是高角羚属的唯一物种,下有高角羚指名亚种和黑面高角羚亚种。尽管在形态上很相似,这两个亚种间存在显著的遗传距离,并且两个亚种间没有杂交种被发现。

A. m. melampus 高角羚指名亚种,分布于非洲东部和南部,包括北至肯尼亚中部,南至南非,西至安哥拉东南部的广袤区域。
A. m. petersi 黑面高角羚亚种,分布于非洲西南部,在纳米比亚西北部和安哥拉西南部。

高角羚为中等体型、体态纤细的羚,头体长约130厘米。雄性肩高约75-92厘米,体重53-76公斤;雌性肩高70-85厘米,体重40-53公斤。

雄性高角羚

雌性高角羚与幼崽
该物种呈现明显的两性异形:
雌性无角且体型小于雄性。雄性长有细长、弯曲的角,长度45-92厘米。角具强棱,分叉,横截面呈圆形,基部中空。这种拱形结构角在打斗时互锁,帮助雄性在争斗中将对手甩出去,同时也能保护头骨免受伤害。


面部特征包括眼周白色环纹以及浅色的下巴和口鼻部。耳朵长约30厘米,形状较尖,有黑色出现。

高角羚下颚的牙齿排列相当特别,其具有类似原猴的齿梳的结构,用于互相梳理头部和脖子上的皮毛,清除皮外寄生虫。
高角羚利用其类似齿梳的牙齿排列,通过梳理皮毛来清理蜱虫。
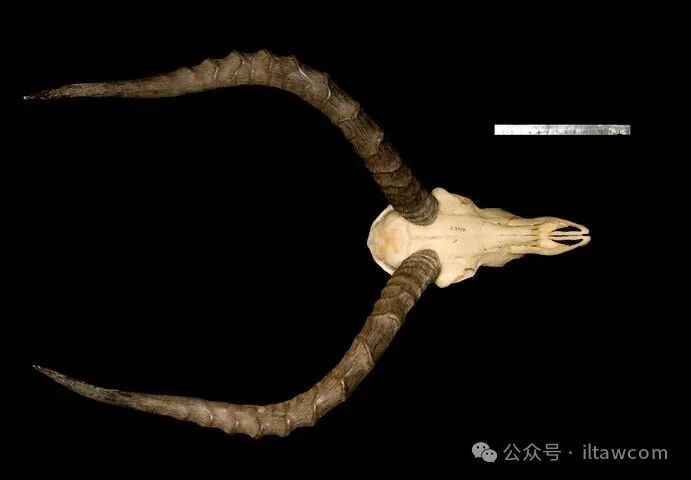

高角羚的骨骼


高角羚其毛皮光滑,呈双色调-红棕色的背部和浅棕色的体侧,与白色的腹部形成鲜明对比。

臀部至后腿上部有黑色条纹,尾巴多毛,长约30厘米,白色为主,带有一道黑色条纹。

高角羚后腿上有被黑色毛簇覆盖的气味腺,其中已鉴定出2-甲基丁酸和2-壬酮等成分。对于有支配地位的雄性,皮脂腺集中于前额和躯干部位,在繁殖季节分泌最为旺盛。对于雌性,皮脂腺则较不发达,活跃程度没有显著的季节性差异。

相比高角羚指名亚种,黑面高角羚亚种体型更大,皮毛颜色更深,这一特征是由隐性基因决定的。


黑面高角羚最显著的特征是鼻子左右各有一条深色条纹,其他特征包括耳朵上更大的黑色尖端,更多毛、更长的尾巴等。

高角羚主要栖息于疏林中,以便利用树荫遮阳,也会在疏林和热带草原间的群落交错区活动,偏好离水源近的地方。栖息地的选择随季节变化:在雨季,高角羚偏好阿拉伯胶树疏林;旱季则偏好镰荚金合欢疏林。另一个影响栖息地选择的因素为是否易受捕食者攻击,高角羚倾向于远离较高的草丛,因为捕食者可能藏在其中。
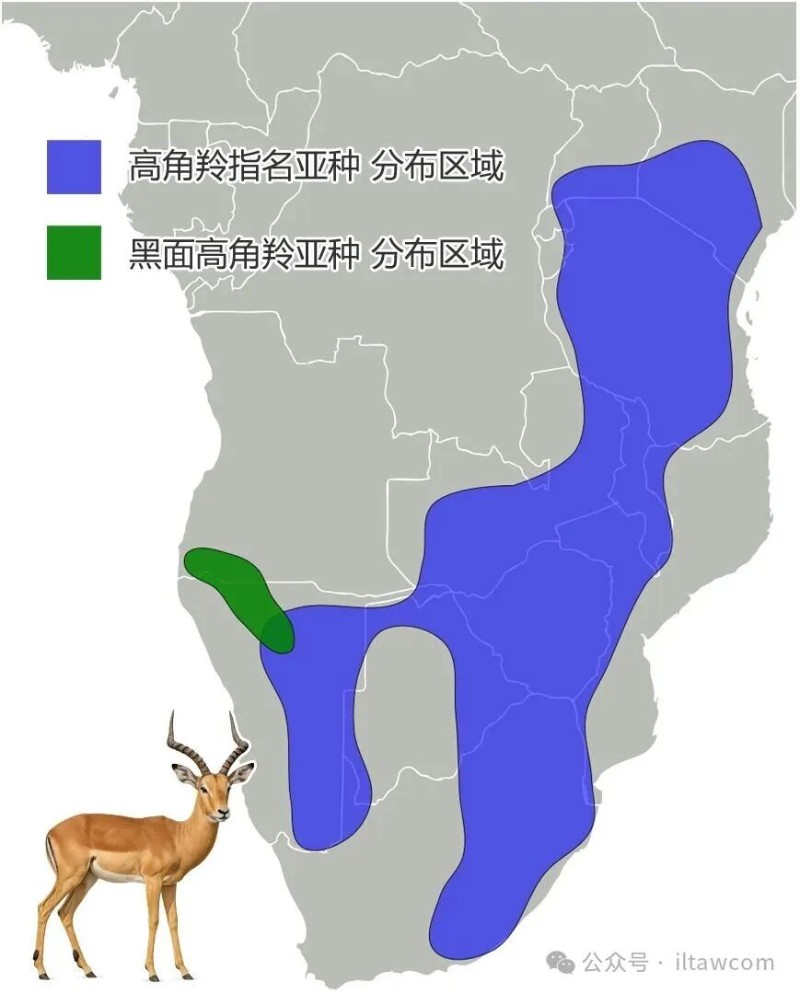
高角羚历史上广泛分布于非洲东部到南部,这一范围很大程度上保持完整,仅在布隆迪等地消失。高角羚生活的范围东至肯尼亚中部和南部、乌干达东北部,南至夸祖鲁-纳塔尔省北部,西至纳米比亚和安哥拉南部,同时被引入加蓬和非洲南部的多个自然保护区。黑面高角羚则限于安哥拉西南部和纳米比亚西北部的卡奥科兰。

高角羚是昼行动物,主要在正午最热的时段以外的昼间时段活动,夜间休息为主。
高角羚既采食灌木嫩枝叶也采食草,何种方式占主导取决于资源的可用性。其食物组成包括单子叶植物、双子叶植物、非禾本科草本植物、果实和金合欢荚果。

高角羚偏好距离水源近的区域,其次偏好多肉植物丰富的区域。一项研究显示,高角羚的食谱包括45%的单子叶植物,45%的双子叶植物,10%的果实;食谱中草的比例在雨季的第一场雨后显著增加,可高达90%,在旱季回落。在雨季结束前后和旱季期间,高角羚多数时候取食高处的嫩叶,特别是在旱季中,高角羚主要取食木本双子叶植物,营养较为缺乏。在不同种类的草中,高角羚主要取食柔软而富有营养的草。

群体进食时,外围的高角羚通常比中心取食的个体对捕食者更警惕;正在进食的个体会采取低头的方式来防备食物被抢占。一项研究显示,觅食时间在旱季末期达到峰值(占白天的75.5%),在雨季期间减少,并在旱季初期达到最低(57.8%)。

高角羚主要存在三种不同的社会群体:有领地意识的雄性、待交配的雄性群体、雌性群体。

有领地意识的雄性会以尿和粪便划分领地的界线,驱赶其他幼年个体或雄性侵入者以守卫领地,并可能在其领地上形成由雌性组成的后宫。
待交配的雄性群体一般比较小,只有不到30只高角羚,两两之间相隔2.5-3米左右,除了年龄差距较大的个体间以外,基本不进行互动。
雌性群体的规模则从6只到100只都有可能,群体在约80-180公顷的范围内活动。高角羚母子间的关系不强,断奶后即告终结,未成年的个体离开母亲的群体,加入其它群体。雌性群体组织较为松散,没有明显的领导。

在待交配的雄性群体和雌性群体中,互相梳理皮毛是一种重要的社交互动。事实上,高角羚是目前已知仅有的既能自己梳理皮毛,又能互相梳理皮毛的有蹄类动物。在互理中,雌性通常梳理有亲缘关系的个体,而雄性则与无亲缘关系的个体互动。高角羚可通过气味腺分泌物识别有领地意识的雄性,其领地范围相对稳定,对于较年长的雄性,领地可以保持数年不变。
高角羚是非洲大型食肉动物,如:猎豹、花豹、野狗、狮子、鬣狗、鳄鱼和蟒蛇,的重要猎物。

高角羚具有其特有的两种跳跃方式:

一种跳跃方式可以跃过高达3米的草木障碍甚至其他高角羚,水平位移可达10米。

另一种跳跃方式由一 连串的跳跃组成,其中每次跳跃均由前腿先落地,后腿在半空中做出踢的动作,四腿落地后再反弹。两种跳跃方式均会跳向不同方向,以迷惑捕食者。有时,高角羚也会隐藏在草木之中,避免被捕食者发现。
高角羚最显著的发声是通过闭合嘴发出一至三声响鼻(呼啸声),随后张开嘴发出二至十声深沉咕噜声,同时下巴和尾巴抬起,这种呼啸声可以传出2千米之远。

高角羚与牛椋鸟存在共生关系,牛椋鸟以高角羚自身无法触及的身体部位(如耳朵、颈部、眼睑、前额和腹部)的蜱虫为食。

一项研究观察到,尽管存在其他动物(如柯氏狷羚、葛氏瞪羚、汤氏瞪羚和转角牛羚),牛椋鸟仍选择性地照料高角羚。一个可能的解释是,由于高角羚栖息于疏林地带(蜱虫密度可能较高),其单位体表面积的蜱虫生物量可能更大。

普通非洲象的活动会对高角羚的数量有正面影响:普通非洲象的活动会造成疏林的减少,并且促进山柑属等灌木丛形成,从而在旱季为高角羚提供了更多食物。

繁殖期的雄性通过发出响亮的咆哮和相互追逐来争夺支配地位,它们步态僵硬,并展示其颈部和角。雄性在繁殖期停止取食和互理行为,可能是为了投入更多时间争取发情雌性。雄性通过检查雌性的尿液来确认其是否处于发情期。遇到此类雌性时,兴奋的雄性开始求偶。

雌性妊娠期持续六至七个月,如果环境过于恶劣,生产的日期可能推迟一个月。高角羚通常为单胎,刚出生的高角羚会被隐藏保护若干周,然后加入群体中的保育小组。高角羚幼崽接受哺乳四至六个月,之后年轻的雄性会被赶出群体,加入待交配的雄性群体;年轻的雌性则可以继续留在本群体中。
雄性一岁性成熟,但成功交配通常发生在四岁后。成年雄性开始建立领地并试图接触雌性。雌性一岁半后可受孕。

世界自然保护联盟(IUCN)将高角羚整体评估为无危物种。然而,黑面高角羚亚种被评估为易危,截至2008年,估计野外存活数量少于1000头,盗猎和自然灾害已显著导致此亚种的衰退。

高角羚指名亚种的种群数量估计约为两百万头,其中约四分之一分布于保护区内,如奥卡万戈三角洲(博茨瓦纳);马赛马拉和卡贾多(肯尼亚);克鲁格国家公园(南非);鲁阿哈和塞伦盖蒂国家公园及塞卢斯禁猎区(坦桑尼亚);卢安瓜河谷(赞比亚);万基国家公园和赞比西河谷(津巴布韦)。